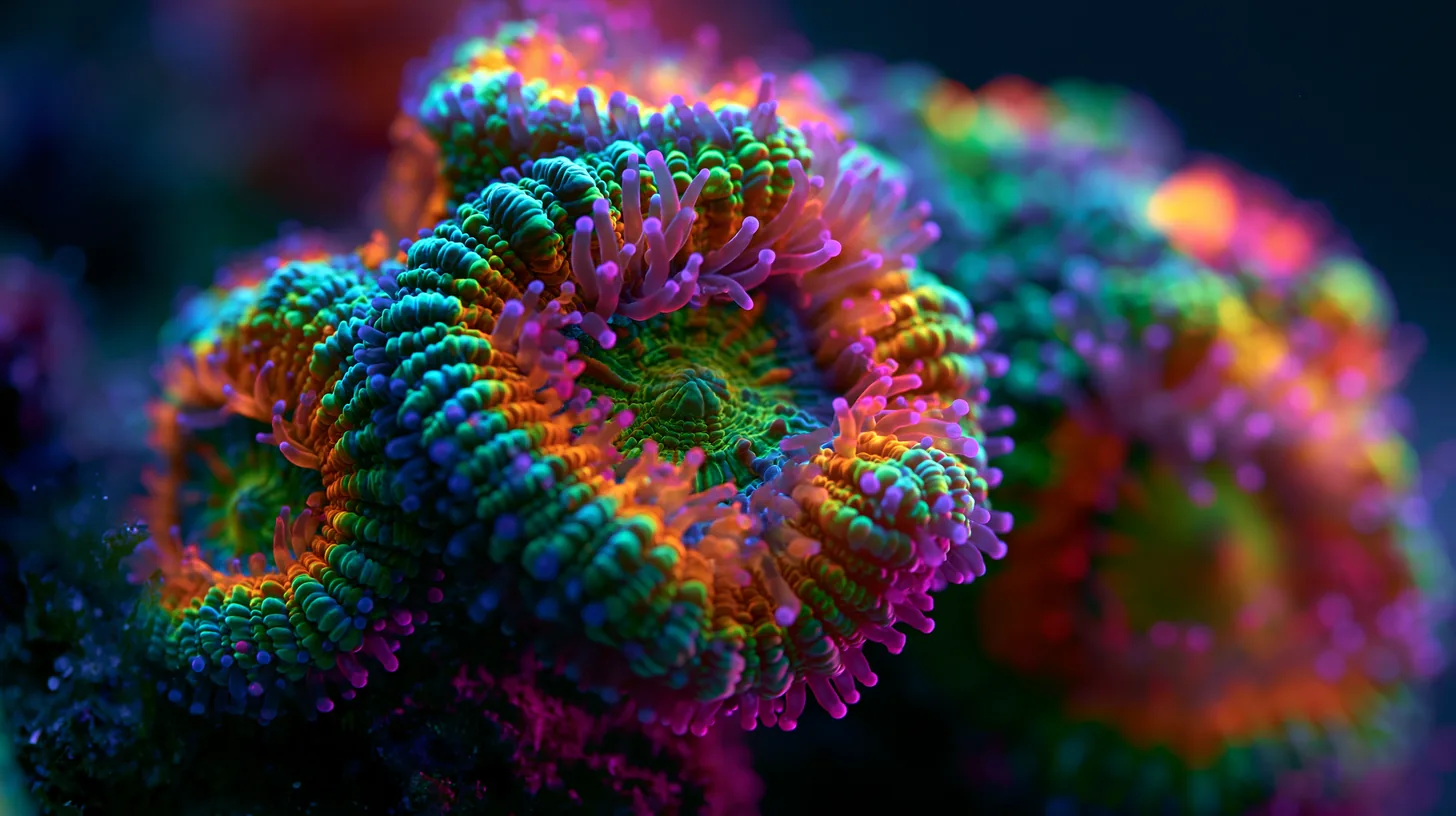
Korallien väri ja pigmentit — GFP-proteiineista zooxanthellaedensiteettiin
Korallien värit ovat yksi riuttaharrastuksen visuaalisista huippuhetkistä — mutta ne ovat myös ikkuna korallien fysiologiaan. Väri ei ole sattumanvarainen esteettinen ominaisuus: se on biologinen signaali, joka kertoo valotilasta, ravinnetilasta ja stressitasosta. Kun ymmärtää, miten väri syntyy ja miksi se muuttuu, saa käyttöönsä diagnostisen työkalun, jota mikään mittari ei korvaa.
Kaksi värinlähdettä
Korallissa on kaksi toisistaan riippumatonta värinlähdettä, jotka toimivat eri mekanismeilla ja reagoivat eri olosuhteisiin.
Zooxanthellaet — korallien sisällä elävät endosymbionttidinoflagellaatit (Symbiodiniaceae) — ovat luonnostaan kellertäviä tai ruskeita. Ne sisältävät klorofylliä, karotenoideja ja peridiniini-klorofylli-a-proteiini(PCP)-komplekseja, jotka absorboivat valoa fotosynteesiä varten. Kun zooxanthellaedensiteetti on korkea, niiden ruskea väri peittää alleen muut pigmentit. Mitä enemmän korallissa on zooxanthellaeja, sitä ruskempana se näyttäytyy — riippumatta siitä, mitä muita pigmenttejä sen kudoksissa on.
Korallien omat pigmentit muodostavat toisen järjestelmän. Ne ovat GFP-superfamilian (green fluorescent protein) proteiineja — siis korallien itsensä tuottamia, eivät symbionttien. Tähän perheeseen kuuluvat sekä fluoresoivat proteiinit (FP) että ei-fluoresoivat kromaproteiinit (CP). Ne voivat tuottaa sinistä, syaania, vihreää, keltaista, oranssia, punaista tai violettia väriä — täysin riippumatta zooxanthellaedensiteetistä.
Nämä kaksi järjestelmää toimivat päällekkäin. Koralli voi olla täynnä zooxanthellaeja ja silti kirkkaan sininen, jos se tuottaa runsaasti sinisiä kromaproteiineja. Se voi myös näyttää vihreältä siksi, että sillä on vähän zooxanthellaeja ja paljon vihreää GFP:tä — tai siksi, että sen zooxantellaedensiteetti on korkea mutta GFP ylittää ruskean vaikutuksen. Pelkkä silmämääräinen arvio ei kerro kummasta järjestelmästä väri kulloinkin syntyy.
GFP-superfamilia — proteiinirakenne ja monimuotoisuus
Vihreä fluoresoiva proteiini löydettiin alun perin meduusasta (Aequorea victoria) vuonna 1962. Cnidaria-eläimillä — johon korallit kuuluvat — on evoluution myötä syntynyt laaja GFP-homologien kirjo, jonka optinen monimuotoisuus ylittää alkuperäisen löydön moninkertaisesti.
Kaikki GFP-superfamilian jäsenet jakavat saman rakenteellisen perustan: β-tynnyri, jonka sisällä oleva tripeptidikromofori absorboi ja (fluoresoivissa tapauksissa) emittoi valoa. Vaikka rakenne on konservoitunut, kromoforin kemiallinen ympäristö vaihtelee suuresti — ja tämä vaihtelu selittää koko spektrin sinisestä infrapunaan.
Fluoresoivat proteiinit (FP)
Fluoresoivat proteiinit absorboivat valoa ja emittoivat sen pidemmällä aallonpituudella — tätä kutsutaan fluoresenssiksi. Korallien tärkeimmät FP-ryhmät ovat:
Syaanit (CFP, ~480 nm emissio) esiintyvät monilla matalavesilajilla ja osallistuvat valon siroamiseen kudoksessa.
Vihreät (GFP, ~510 nm emissio) ovat yleisin ryhmä. Ne absorboivat sinistä valoa (~488 nm) ja emittoivat vihreänä. GFP:n ilmentyminen on valosta riippuvainen: sininen valo indusoi tuotantoa matalasyvyisissä korallissa. Bollati ym. (2022, eLife) osoittivat, että GFP-proteiinit hienosäätävät korallien sisäistä valoympäristöä moduloimalla säteilyä kudoksen läpi.
Punaiset (RFP, ~580–620 nm emissio) tuottavat korallien punaiset, oranssit ja roosat sävyt. Erityinen alaryhmä, fotokonvertoituvat punaiset fluoresoivat proteiinit (pcRFP), ovat biologisesti erityisen merkittäviä: ne syntetisoituvat ensin vihreinä, mutta UV-valo (~390 nm) konvertoi kromoforin pysyvästi punaiseksi emittoivaksi muodoksi. Konvertoituneet pcRFP:t absorboivat sinivihreää valoa ja emittoivat oranssinpunaisena — aallonpituuskonversio, jolla on kriittinen merkitys syvemmissä elinympäristöissä.
Kromaproteiinit (CP)
Kromaproteiinit ovat GFP-homologeja, jotka absorboivat valoa mutta eivät emittoi sitä merkittävästi — ne siis “nielevät” valon ilman fluoresenssia. Tämä tekee niistä tehokkaita valon vaimentimia. CP:t tuottavat korallien violetit, tumman punaiset, ruskeat ja mustat sävyt. Useimmat CP:t absorboivat voimakkaimmin oranssissa-keltaisessa alueella (562–586 nm).
Biologiset tehtävät: mitä pigmentit tekevät?
GFP-superfamilian proteiinit eivät ole passiivisia väriaineta. Ne ovat funktionaalisia molekyylejä, joilla on aktiivisia biologisia tehtäviä. Tutkimus on tunnistanut kolme päämekanismia, jotka eivät sulje toisiaan pois.
1. Fotoprotektio matalilla riutoilla
Matalilla riutoilla korallit altistuvat valonylitarjonnalle — PAR-arvot voivat ylittää 3900 µmol m⁻² s⁻¹ aaltojen linssivaikutuksen seurauksena (Wangpraseurt ym. 2014). Ylikuormitus vahingoittaa zooxanthellaeja: superoksidia ja muita reaktiivisia happiyhdisteitä (ROS) syntyy, klorofyllin fotoinhibitio käynnistyy.
CP:t toimivat tässä yhteydessä biologisena aurinkosuojana. Ne sijaitsevat ektodermissa, zooxanthellaeja sisältävän endodermikerroksen päällä. Absorptiomaksimissaan (562–586 nm) ne voivat vähentää klorofyllin eksitaatiota jopa 50 % (Bollati ym. 2022). Tämä on erityisen tärkeää bleachingin aikana: kun zooxanthellaedensiteetti laskee, korallien luustoheijastus ja kudoshajonta lisäävät valoa kudoksessa — CP:t nousevat hillitsemään tätä valovahvistusta.
GFP:t poistavat myös reaktiivisia happiyhdisteitä suoraan — tämä antioksidanttifunktio on erillinen fotoprotektiivisesta roolista.
2. Aallonpituuskonversio syvissä ympäristöissä
Mesofotiset riutat (50–150 m syvyys) ovat valon suhteen täysin erilaisessa tilanteessa. UV ja pitkät aallonpituudet vaimenevat nopeasti, jolloin valo on lähes yksinomaan sinivihreää. Punainen valo, jota klorofylli hyödyntää tehokkaasti syvemmissä kudoskerroksissa, on käytännössä poissa.
Tähän pcRFP:t tuovat ratkaisun. Bollati ym. (2022) osoittivat suorilla optomikrosensorimittauksilla, että konvertoituneet pcRFP:t voivat kaksinkertaistaa punaisen säteilyn (560–650 nm) määrän 50–80 metrin syvyydessä — ja yli 80 metrissä pcRFP-fluoresenssi on dominoiva oranssipunaisen valon lähde zooxanthellaeja ympäröivässä kudoksessa. Oranssi-punainen valo läpäisee syvemmät kudoskerrokset paremmin kuin sinivihreä, koska symbionttipigmentit absorboivat sitä vähemmän. Tulos: pcRFP-korallit pärjäävät syvemmissä valotiloissa kuin ilman niitä elävät lajitoverit.
3. Aktiivinen kasvualueiden ja palautumisen tuki
GFP-tyyppiset proteiinit kertyvät voimakkaasti alueisiin, joissa zooxanthellaetiheys on matala: kasvavat latvat, haarautuvien korallien kärjet, mekaanisista vaurioista toipuvat alueet. Bollati ym. (2020) ehdottivat, että tämä on tarkoituksellinen optinen palautesilmukka: matala symbionttikonsentraatio johtaa lisääntyneeseen valonläpäisyyn kudoksessa → valo indusoi GFP/CP-tuotantoa → pigmentit tasoittavat valokliimaa → uudet symbionttipopulaatiot pystyvät asettumaan ilman fotodamage-riskiä.
Valon vaikutus värin intensiteettiin
GFP-proteiinien ilmentyminen on valoherkkä prosessi, jonka laukaisumekanismina toimii sininen valo. D’Angelo ym. (2008) osoittivat, että GFP-proteiineilla on valoindusoitu transkriptionaalinen säätely: sininen valo aktivoi proteiinin tuotantoa, pimeys hillitsee sitä.
Käytännön seuraus: koralli, joka siirretään korkeampaan PAR-ympäristöön, kasvattaa GFP/CP-tasojaan muutamien viikkojen kuluessa. Tämä on selvä havaittava ilmiö: korallin väri voimistuu siirron jälkeen — ei siksi, että koralli “parantuisi”, vaan siksi, että se tuottaa enemmän suojaavia pigmenttejä vasteena lisääntyneelle valopaineelle.
Kääntäen: koralli matalassa valossa tuottaa vähemmän valoindusoituneita GFP:itä. Vähävaloisessa altaassa pidetty teoriassa kirkkaan sininen laji voi näyttää tummemmalta — ei sairauden merkkinä, vaan sopeutuneena vasteena.
Tärkeä poikkeus: syvien lajien pcRFP:t ovat konstitutiivisesti ilmentyviä — ne eivät reagoi valotasoon samalla tavalla. Ne on “ohjelmoitu” pysymään korkeina riippumatta valon intensiteetistä, koska syvissä ympäristöissä ei ole ylivalotusriskiä jonka varalta ne pitäisi reguloida alaspäin.
Ravinteiden vaikutus väriin: zooxanthellaedensiteetti himmentimenä
Ravinteilla on kokonaan eri mekanismi värin säätelyssä kuin valolla — ne eivät säätele GFP-tuotantoa suoraan, vaan ohjaavat zooxanthellaedensiteettiä, joka puolestaan peittelee tai paljastaa alla olevat pigmentit.
Korkeat nitraatti- ja fosfaattitasot lannoittavat zooxanthellaeja. Kun symbionttimäärä kasvaa, niiden ruskea väri peittää alleen FP:t ja CP:t. Korallit ruskettuvat. Tämä ei ole merkki terveydestä — se on merkki siitä, että symbioosisuhde on vinoutunut: symbiontti hyötyy ylenpalttisista ravinteista, koralli ei saa niiden suorituksesta lisähyötyä.
Liian matalat ravinnetasot puolestaan johtavat zooxanthellaepopulaatioiden heikkenemiseen. Ensimmäisenä koralli vaalenee — vähemmän ruskeaa peittämässä alta. Sitten tulee varsinainen bleaching, kun symbionteita karkotetaan aktiivisesti. Äärimmäisessä bleachingissa koralli on valkoinen tai kirkkaan vaaleanvihreä (jos GFP:tä on paljon ilman zooxanthellaekattoa).
Käytännössä tämä tarkoittaa, että kirkkaat värit eivät ole merkki ravinteiden puutteesta — ne voivat olla merkki siitä, että zooxanthellaemäärä on optimaalinen eikä ylimääräistä symbionttien rusketta ole peittämässä. Suositeltavat ravinnetasot, joissa koralliväri usein on parhaimmillaan: nitraatti 2–10 ppm, fosfaatti 0,03–0,08 ppm.
Väri bleachingissa — optinen palautesilmukka
Bleachingissa väridynamiikka muuttuu dramaattisesti. Kun zooxanthellaet karkotetaan lämpöstressin, valoylitarjonnan tai muun stressin takia, kaksi asiaa tapahtuu samanaikaisesti:
- Ruskea symbionttipeite katoaa — alla olevat FP:t ja CP:t paljastuvat.
- Zooxanthellaiden puuttuessa kudosvaloympäristö kirkastuu: luuston heijastus ja kudoshajonta vahvistavat valon intensiteettiä kudoksen sisällä.
Bollati ym. (2022) osoittivat, että tämä lisääntynyt sisäinen valo aktivoi CP-tuotantoa sinivalon kautta: valoistunut kudos indusoi lisää kromaproteiineja. Bleachingin aikana koralli voi siis paradoksaalisesti muuttua kirkkaamman pinkkiä tai violettia stressireaktion aikana — tämä on tunnettu “pinkki koralli” -ilmiö, ei tervehtymismerkki vaan fotoprotektiivinen vastesilmukka kriisitilanteessa.
Bleachingin läpikäynyt koralli, joka on vaalean sininen tai vaalean vihreä (ei valkoinen), on tyypillisesti säilyttänyt GFP:nsä vaikka zooxanthellaet ovat kadonneet. Pelkästään valkoinen koralli on menettänyt molemmat. Pelkistä pigmenteistä väriään pitävä koralli voi toipua — se on aktiivisesti säätelevä järjestelmä. Täysin valkoinen koralli on kuollut tai erittäin lähellä kuolemaa.
Käytännön sovellukset akvaristille
Väri diagnostisena signaalina. Äkillinen ruskettuminen viittaa joko ravinnepiikin aiheuttamaan zooxanthellaexpansioon tai valotason laskuun. Voimistuva väri korkean valon jälkeen on normaali fotoadaptaatiovaste. Bleaching on hätäsignaali — selvitä lämpötila, valo ja ravinteet ennen kuin teet korjausliikkeitä.
Sinivalo paljastaa, täysspektri rakentaa. Fluoresoivat proteiinit aktivoituvat parhaiten sopivilla aallonpituuksilla — usein sinisellä tai UV:llä — ja niiden fluoresenssi näyttää dramaattisimmalta sinivalossa. Tästä tulee usein harhaluulo: “sininen valo tekee värit”. Todellisuudessa täysspektri päivällä rakentaa pigmentit (indusoi GFP:tä), sinivalo illalla vain paljastaa ne fluoresenssina. Aja täysspektriä huippufotoperiodilla.
Kirkas väri ei ole merkki stressistä. Voimakas fluoresenssi — erityisesti sininen, vihreä tai syaani — voi merkitä, että koralli on tuottanut runsaasti valoindusoituneita pigmenttejä vasteena korkealle valolle. Tämä on normaali biologinen sopeutuminen, ei sairauden merkki. Huolehdi ravinnetasapainosta ja valon tasaisuudesta — väriä ei tarvitse erikseen “edistää”.
Lähdeluettelo
1. Vertaisarvioidut tutkimukset
- Bollati, E. et al. (2022). Green fluorescent protein-like pigments optimise the internal light environment in symbiotic reef-building corals. eLife, 11, e73521. https://doi.org/10.7554/eLife.73521
- Bollati, E. et al. (2020). Functional colour: GFP-like proteins in the coral Acropora millepora during bleaching. Proceedings of the Royal Society B, 287, 20200626.
- D’Angelo, C. et al. (2008). Blue light regulation of host pigment in reef-building corals. Marine Ecology Progress Series, 364, 97–106.
- D’Angelo, C., Denzel, A., Vogt, A., Matz, M.V., Oswald, F., Salih, A. et al. (2008). Blue light regulation of host pigment in reef-building corals. Marine Ecology Progress Series, 364, 97–106.
- Salih, A. et al. (2000). Fluorescent pigments in corals are photoprotective. Nature, 408, 850–853. https://doi.org/10.1038/35048564
- Smith, E.G. et al. (2013). Screening by coral green fluorescent protein (GFP)-like chromoproteins supports a role in photoprotection of zooxanthellae. Coral Reefs, 32, 463–474. https://doi.org/10.1007/s00338-012-0994-9
- Smith, E.G. et al. (2017). Acclimatization of symbiotic corals to mesophotic light environments through wavelength transformation by fluorescent protein pigments. Proceedings of the Royal Society B, 284, 20170320. https://doi.org/10.1098/rspb.2017.0320
- Alieva, N.O. et al. (2008). Diversity and Evolution of Coral Fluorescent Proteins. PLOS ONE, 3(7), e2680. https://doi.org/10.1371/journal.pone.0002680
- Wangpraseurt, D. et al. (2014). In vivo microscale measurements of light and photosynthesis during coral bleaching. Journal of Experimental Biology, 217, 489–498.
- DiPerna, S. et al. (2018). Effects of variability in daily light integrals on the photophysiology of the corals Pachyseris speciosa and Acropora millepora. PLOS ONE, 13(9), e0203882.
2. Harrastajakirjallisuus ja -lähteet
- Aslett, C.G. (2024). The Complete Reef Aquarist. Reef Ranch Publishing Ltd.
- Riddle, D. (2007). Coral Coloration: Proteins and Chromoproteins. Reefs.com.